The Amarasekare Lab
UCLA Department of Ecology and Evolutionary Biology
Research
My research focuses on how the interplay between abiotic environmental variation and biotic interactions influences ecological and evolutionary dynamics. It seeks to explain patterns of dynamics and diversity that are observed in nature, and to predict how such patterns may change under perturbations to the abiotic and biotic environment. The distinctive features of my work are the strong mechanistic focus and the tight integration between theory and data. My work is also broad, spanning investigations of the interplay between biotic interactions and temporal variation on the one hand, with temperature as the axis of abiotic variation, and biotic interactions and spatial variation on the other, with dispersal as a mechanism for sampling spatial heterogeneity.
My research contributions fall into three main areas:
- 1. Spatial dynamics of population regulation, species interactions and the evolution of dispersal
- 2. Dynamics and diversity of multi-trophic communities
- 3. Effects of temperature variation on population dynamics, species diversity and the evolution of thermal reaction norms
1. Spatial dynamics of population regulation, species interactions and the evolution of dispersal
 1.1 Spatial dynamics of multi-trophic communities
1.1 Spatial dynamics of multi-trophic communities
My research has made an important contribution towards developing spatial theory for single species populations (Amarasekare 1998a, 1998b, Amarasekare 2004c}, competitive communities (Amarasekare 2004b, 2004d, Shurin et al. 2004, Frank and Amarasekare 1998}, mutualistic interactions (Amarasekare 2004a} and marine communities subject to harvesting (Micheli et al. 2004). This body of work has resulted in several influential syntheses (Amarasekare 2003b, Leibold et al. 2004, Ashman et al. 2004, Chase et al. 05, Mouquet et al. 2005, Amarasekare 2008c, Urban et al. 2008}. Below I highlight two major research contributions I have made since then.
I have developed a body of theory on the spatial dynamics of multi-trophic communities (Amarasekare 2006, Amarasekare 2007b, Amarasekare 2008b, Amarasekare 2009a, Amarasekare 2010), which illustrates the novel emergent properties arising from the interplay between competition, consumer-resource interactions and dispersal. Importantly, the dispersal strategy and behavior of an inferior competitor that is less susceptible to predation can have a strong effect on a superior competitor that is more susceptible to predation, but the latter species' dispersal traits have no effect on the former. The key insight is that dispersal asymmetries arising from trophic constraints can create keystone dispersers, species whose dispersal has a disproportionately large effect on community dynamics. My work shows that such species can have a significant impact on both diversity maintenance and the control of invasive pests.
1.2 Evolution of dispersal in multi-trophic communities
My work (Amarasekare 2016a) is the first to investigate how the tension between resource acquisition and predator avoidance influences the evolution of dispersal in multi-trophic communities. I find that dispersal evolution is possible even when the consumer species is not directly affected by environmental variability, but rather experiences the consequences that such variability has on its resource and predator. This finding underscores the importance of indirect ecological interactions in shaping the evolution of dispersal. While the ecological community can provide a strong selective environment for the evolution of dispersal, the nature of interactions between trophic levels can also impose constraints on evolution. Ongoing work investigates how non-random dispersal strategies influence the establishment of invasive species (Beckett and Amarasekare, In Prep.), the co-evolution of dispersal and resource/consumer traits in multi-trophic communities (Fronhofer and Amarasekare, In Prep.), and how temperature-dependent dispersal influences species' range shifts in response to climate warming (Amarasekare 2016b).
Example papers:
2. Dynamics and diversity of multi-trophic communities
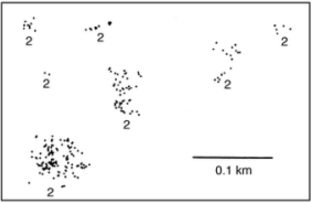
 My empirical work on an insect host-parasitoid system in southern California Amarasekare (2000a, Amarasekare 2000b, Amarasekare 2003a) has inspired the development of theory on how the interplay between local non-linear dynamics (driven by intraguild predation) and temporal environmental variation influence species coexistence in multi-trophic communities (Amarasekare 2003a, Amarasekare 2007a, Amarasekare 2008a). The hallmark of this work is the tight integration between theory and data: theory inspired by the host-parasitoid system generated predictions that were tested with laboratory and field experiments; the experiments in turn provided insights for developing more biologically realistic theory. This work has also yielded general insights into the dynamics and diversity of multi-trophic communities, two of which I highlight below.
My empirical work on an insect host-parasitoid system in southern California Amarasekare (2000a, Amarasekare 2000b, Amarasekare 2003a) has inspired the development of theory on how the interplay between local non-linear dynamics (driven by intraguild predation) and temporal environmental variation influence species coexistence in multi-trophic communities (Amarasekare 2003a, Amarasekare 2007a, Amarasekare 2008a). The hallmark of this work is the tight integration between theory and data: theory inspired by the host-parasitoid system generated predictions that were tested with laboratory and field experiments; the experiments in turn provided insights for developing more biologically realistic theory. This work has also yielded general insights into the dynamics and diversity of multi-trophic communities, two of which I highlight below.
2.1 Transient dynamics in prey-natural enemy interactions and biological pest control
There is increasing recognition of the importance of transient dynamics in ecological interactions, but very little is known about their role in biological pest control. Using a combination of mathematical models and manipulative experiments, we have shown that weak self-limitation, a characteristic of many pests that attack crop monocultures, combined with the saturating functional responses of natural enemies, can lead to strong transient fluctuations that cause the short-term dynamics to be dominated by pest outbreaks even when the long-term outcome is a stable pest-enemy equilibrium (Kidd and Amarasekare 2012}. Release of multiple enemy species can compromise pest suppression if interactions between enemies lead to stronger transient fluctuations than when a single enemy attacks the pest. Intra-specific interference, long thought to be an undesirable property in a biocontrol agent, can improve short-term pest control by reducing the magnitude and duration of transient fluctuations. These findings underscore the key role that transient dynamics play in pest-enemy interactions. The tendency to induce strong transient fluctuations (e.g., through a long handling time) is therefore a crucial consideration when identifying effective pest control agents. This work was highlighted as an In Focus article in the January 2012 issue of the Journal of Animal Ecology.
2.2 The role of handling time in generating consumer-resource oscillations
Interaction strength, quantified as the attack rate, has long been considered the metric for predicting consumer-resource oscillations, despite the fact that it is the handling time that induces the positive feedback responsible for generating oscillations. We have recently shown (Johnson and amarasekare 2015) that it is the multiplicative effect of the attack rate and handling time that mediates the positive feedback, and that oscillatory tendency is determined by the strength of this effect relative to the negative feedback generated by resource and/or consumer self-limitation. Tests of model predictions with data reveal that the majority (77%) of consumer-resource interactions involve low attack rates and handling times, requiring only a modest level of self-limitation to suppress oscillations. Only 23% of the interactions exhibit a strong oscillatory tendency, consistent with previous findings, based on time-series data, that 30% of consumer-resource interactions in nature exhibit oscillations. Our findings are highlighted in a recent article in TREE (Borelli et al. 2015} on stability selection across ecological scales. Inspired by these findings, we are now investigating the coevolution of attack rates and handling times in the face of population-level constraints imposed by stochastic extinction of consumer-resource interactions with a high oscillatory tendency (Johnson and Amarasekare, In Prep.).
Example Papers
3. Effects of temperature variation on ecological and evolutionary dynamics
A major achievement in recent years is the development of a mechanistic theoretical framework for elucidating (i) how temperature effects on life history (reproduction, development, survivorship) and interaction traits (competition coefficients, attack rates, handling times) influence fitness, population regulation and species coexistence, and (ii) how these ecological processes feed back into trait responses, causing trait evolution.
 3.1 From traits to demography: temperature effects on fitness
3.1 From traits to demography: temperature effects on fitness
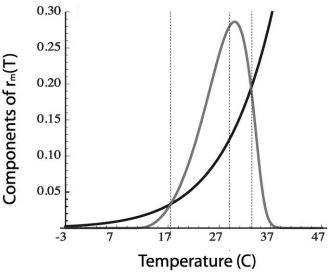
My recent work (Amarasekare and Savage 2012, Amarasekare and Sifuentes 2012} illustrates the power of the trait-based approach for elucidating temperature effects on demographic processes. We used age-structured population dynamics to derive an analytical expression for the temperature dependence of fitness in terms of the temperature responses of fitness components (fecundity, development, survivorship), which provides exact calculations of the thermal limits to viability and the optimal temperature for fitness for any multicellular ectotherm species. Our framework thus provides a rigorous, and general, method for quantifying extinction risk due to climate warming.
3.2 From traits to populations: temperature effects on non-linear dynamics
The theory we have developed using delay differential equations (DDEs) (Amarasekare and Coutinho 2013, Amarasekare and Coutinho 2014, Johnson et al. 2015) constitutes a major breakthrough in analyzing temperature effects on population dynamics. It explicitly incorporates temperature-dependent developmental delays and mechanistic descriptions of trait responses to temperature, the first time these biological features have been integrated into a common mathematical framework. Our theory makes testable comparative predictions about how the interplay between temperature and intra-specific competition influences life history traits and how these trait responses translate into population dynamics. A key insight is that warming effects on population dynamics depends crucially on the mechanism by which temperature affects self-limitation. When competition strength increases with temperature, warming increases the risk of stochastic extinction by lowering minimum population sizes; when competition is strongest at temperatures optimal for reproduction, warming increases the potential for population outbreaks by increasing the maximum population size. These results underscore the counter-intuitive outcomes that ensue when perturbations in the abiotic environment percolate through the non-linear feedback loops that underlie biotic interactions.
3.3 From traits to communities: temperature effects on species interactions
In my most recent work I develop a trait-based framework for investigating temperature effects on species interactions. Here I highlight two major accomplishments on this front.
I have developed theory to investigate temperature effects on the negative and positive feedback processes that underlie consumer-resource interactions. A model that incorporates, for the first time, the empirically observed non-monotonic temperature responses of reproductive and consumptive traits yields radically different predictions about warming effects compared to previous studies that consider only monotonic responses (Amarasekare 2015). Contrary to previous findings, warming does not always amplify consumer-resource oscillations. In fact, warming can dampen oscillations depending how temperature affects self-limitation. Moreover, whether warming impacts diversity through deterministic or stochastic extinction depends on whether the consumer is a thermal generalist or specialist compared to the resource. These results highlight the critical importance of investigating temperature effects on both negative and positive feedback processes that underlie consumer-resource interactions.
The second contribution involves elucidating the mechanisms by which climate warming causes phenological shifts. Such shifts are the direct result of temperature effects on development: warmer temperatures lead to faster development and earlier emergence. We develop DDE models with temperature-dependent developmental delays to investigate (i) how temperature responses of life history traits translate into phenological shifts at the population level and (ii) how species-specific phenological shifts disrupt species interactions (Scranton and Amarasekare, In Prep). A novel, and unexpected, finding is that phenological shifts are driven by the multiplicative effect of the temperature responses of development and juvenile mortality}, which allows us to predict the exact magnitude of the shift based solely on knowledge of how temperature affects the underlying traits.
3.4 Evolution of thermal reaction norms
A complete understanding of temperature effects on species interactions requires elucidating the feedbacks between ecological dynamics and reaction norm evolution. I address this issue using a combination of empirical and theoretical approaches. On the theoretical front I have developed a framework for investigating the evolution of thermal reaction norms that integrates, for the first time, quantitative genetic models of trait evolution with stage-structured models of population dynamics. I use this framework to explain a recurrent latitudinal pattern in thermal reaction norms: the temperature at which performance is maximized coincides with the mean habitat temperature in tropical ectotherms but exceeds the mean in temperate ectotherms. Our eco-evolutionary model shows that it is the magnitude of seasonal fluctuations and their effect on stage-structured population dynamics that determine the relationship between thermal optima and the mean habitat temperature (Amarasekare and Johnson In Revision). A key finding is a potential directionality in invasion success. Invasive species of tropical origin are more likely to adapt to temperate climates but invasive species of temperate origin are less likely to adapt to tropical climates. On the empirical front, we are quantifying genetic variation in the thermal reaction norms of the harlequin bug and its parasitoids. Preliminary findings on the bugs are fascinating. Development, a trait under strong biochemical control, exhibits no genetic variation in the reaction norm under typical temperatures but offers a hint of hidden variation at higher temperatures; survivorship, a trait affected by resource and natural enemy variability, exhibits considerable genetic variation even under typical temperatures. These findings provide an intriguing glimpse into the genetic landscape underlying the evolution of thermal reaction norms. We are conducting full-scale experiments on the bugs and parasitoids to elucidate, for the first time, how reaction norms coevolve in a natural community.
Example Papers